文 | 追问nextquestion
暴雨倾盆,霸王龙紧逼颤抖的吉普,后视镜闪过猩红眼眸;厨房内,迅猛龙利爪击打瓷砖,角落的人们屏气凝神……《侏罗纪公园》中那些令人心跳加速的追逐场景,让我们深思:
若不是那颗改变地球命运的陨石,或是那场吞噬一切的冰河世纪,面对这些智力与力量都强得惊人的庞然大物们,像人类这样弱小的哺乳动物,真的能在这片丛林中立足吗?
然而,历史总是充满戏剧性的转折。人类登上舞台,与巨兽争锋靠的并非蛮力,而是“运用工具”的独特智慧。正是这份智慧,让我们从弱小的生存者进化为地球的主宰,甚至有希望实现一个令人震撼的构想:让早已消逝的巨龙重现人间。
▷图1.Deep Time Cognition团队主页. 图源:Deep Time Cognition官网
除了恐龙迷,还有一群科研人员也对人类与恐龙之间的关系感兴趣。他们来自一个由认知动物学家、神经解剖学家和古生物学家组成的研究团队DeepTimeCognition,致力于从进化视角探索认知的发展过程。

▷ Osvath M, Němec P, Brusatte SL, Witmer LM. Thought for food: the endothermic brain hypothesis. Trends Cogn Sci. 2024;28(11):998-1010. doi:10.1016/j.tics.2024.08.002.
2024年11月,该团队在期刊Trends in Cognitive Sciences上发表了一项研究,探讨了“全身内温性(whole-body endothermy)”与“意识”演化之间关系。他们注意到恐龙和哺乳动物分别独立演化出了全身内温性特征,与这一变化同时发生的还有神经认知结构的变化:神经元数量激增至原来的20倍,并同时进化出新的大脑结构。研究团队通过充分的证据论证了“全身内温性”与“认知能力发展”之间的深层联系,更展现了从进化生物学和比较生物学角度开展大脑研究的独特视角与意义。
01 全身内温性如何进化得来?
那究竟什么是“全身内温性”呢?这是一种生理特征,使内温性动物(Endotherm)能够通过内部代谢来维持稳定的体温,而不依赖外部环境温度。这种特性需要极高的能量代谢支持,在现存物种中,鸟类和哺乳动物是最典型的代表。在神经认知功能方面,鸟类也与哺乳动物表现出惊人的相似性,而这种神经认知方面相似性在爬行动物中并不存在,尽管从自然进化的角度来看,鸟类与爬行动物更为接近。
可以说,从外温性向内温性的转变,是大脑进化史上最剧烈的变化之一。作为地球上仅存的两类高代谢型内温性动物(tachymetabolic endotherms),鸟类和哺乳动物所共有的这些认知特征,很可能是支撑其高能量代谢生活方式所必需的。这种共性为我们理解认知能力的发展提供了重要线索。

▷图2.简化进化图谱及研究方法论. 图源:Trends Cogn Sci
02 认知与全身内温性有何关系?
“全身内温性”关系到能量消耗和生物代谢,而“认知”则是大脑的核心功能,这两个看似不相关的特征,为何会在进化上紧密关联?仅仅因为“全身内温性”与“认知”同时出现,可能还不足以令人信服。
生物体要自我维持,就需要将外部的能量转化为支持自身细胞活动和结构的能量,这种化学活动网络统称为“代谢”。为了避免崩解,生物体会寻找代谢所需的物质,同时规避潜在的伤害。这种平衡主要通过应变稳态调节(allostasis)来维持,所谓应变稳态调节不仅仅是被动地对变化作出反应,更需要主动预测并应对干扰。而认知,正是这样一个与生物体环境密切相关的稳态调节系统,因此被认为或许与代谢密切相关[1]:
“认知由感官和其他信息处理机制组成,生物体通过这些机制熟悉、评估环境特征并与之进行有效互动,以满足其生存需求。其中最基本的需求,就是生存/存续、成长/繁荣以及繁衍。”
当从这种角度来理解认知,神经系统可以被视为进化上为了维持生物基本的应变稳态的一种延伸,大脑的出现使得生物能够更迅速、更长距离地移动,从而接触到更多信息,并且更好地将这些信息存储在记忆中。
这样看来,大脑在能量摄取中极其重要,那岂不是越发达越好?这样的想法可能忽视了“投资”大脑发育所要付出的代价。“昂贵大脑”假说(the expensive brain hypothesis)认为,当我们拥有一颗更大的大脑时,要么需要增加总能量摄入,要么需要减少其他高能耗的身体活动来补偿。事实上,在能量可用性较低或条件波动较大的环境下,动物通常会出现较小的大脑,也就是说,这些动物会为了减少大脑“支出”而“选择”更小的大脑。
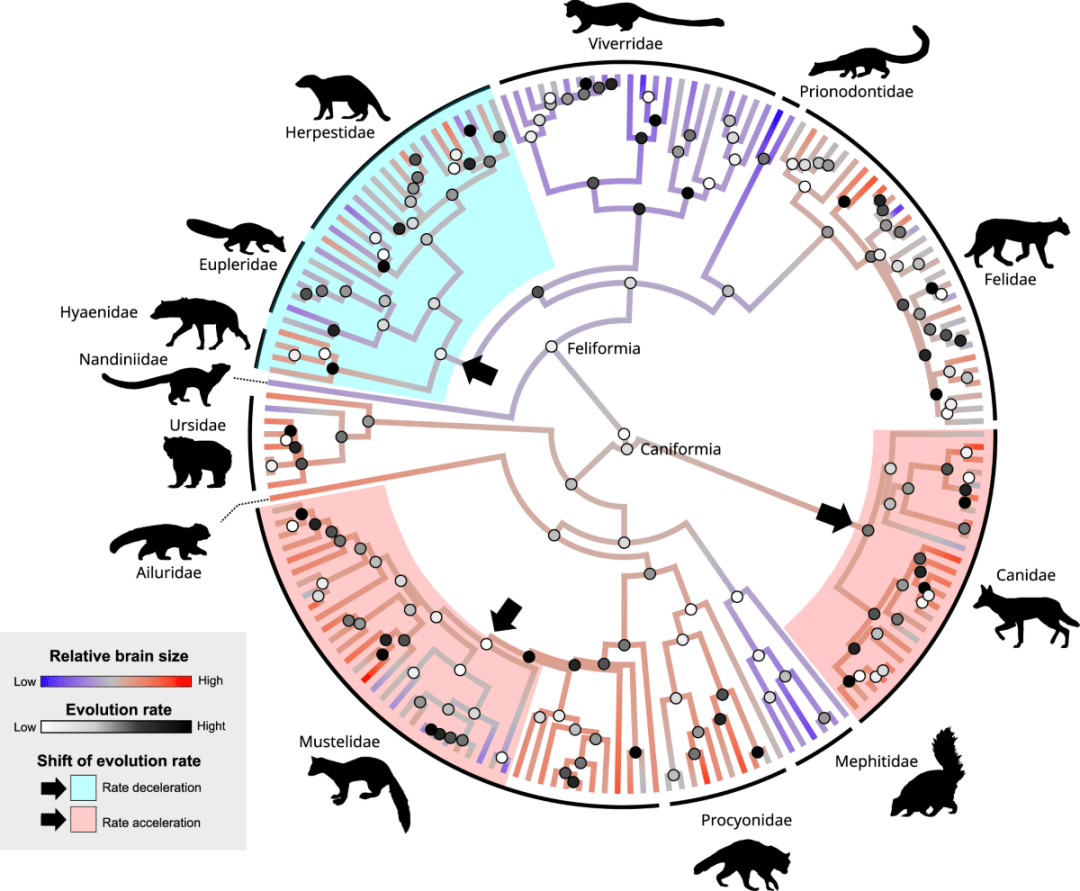
▷图3.食肉动物脑容量演化. 图源:Nature
然而,当比较外温性动物和内温性动物之间的差异时,这种大脑大小的线性变化关系并不成立。在内温性动物中,高耗能的脑神经元数量呈数量级增长——这代表的是神经组织的成本/收益比的变化。一个有意思的观察是:在内温性动物中,拥有较大的大脑往往意味着更长的寿命,而外温性羊膜动物则恰恰相反。这意味着在爬行动物中,对神经组织的投入会分散其他身体活动的能量,加速生命体的衰老。
而内温性从根本上改变了这种关系,大量的高耗能神经元不仅不与其他身体活动竞争,反而为这些活动的维持提供支持。对内温性动物来说,大脑发育是一项高风险但更高回报的“投资”:拥有神经元更丰富、功能更灵活的大脑,其能量代价与大脑本身的功能提升相比微不足道。
更不用说,内温性动物的大脑在抚育后代方面也起到了关键作用。与外温性动物相比,内温性动物的后代抚育行为更为常见且先进。这种现象的本质在于,内温性动物获取的能量,不仅能支持自身的身体活动和大脑运作,还能支持其他个体的需求。
03 大脑进化与能量消耗的博弈,谁赢了?
通过比较内温性鸟类与外温性羊膜动物,我们了解到“全身内温性”和“认知”之间的紧密关系。那么,大脑功能这样的“质的变化”是如何发生的呢?
首先,我们需要知道这两类大脑所支持的认知功能到底存在哪些差异,以及这些差异存在的原因。
当内温性出现时,生命进入了“暴食时代”(the age of gluttony)。在此之前,动物个体从未像内温性动物那样消耗如此大量的食物。现存的内温性动物食物摄入量(按重量计算)是同等体重活跃爬行动物的8倍(哺乳动物)到11倍(鸟类)[2]。
虽然外温性动物也需要食物来维持生命、生长发育和繁殖后代,但内温性动物则还需依赖食物为其高代谢提供能量、产生体热。外温性动物可以在没有食物的情况下依赖外部热能长时间存活,而对食物的较低依赖性让它们还可以通过减少觅食活动来进一步节约能量[3]。

▷图4.冬眠中的箱龟. 图源:Boxturtles.com
基于这些差异,可以推断,内温性动物认知功能的进化主要是为了获取食物。它们的高代谢使其能够更远距离地觅食,并能应对环境温度的波动。然而,高代谢也使得这种高能耗的觅食行为成为必要。
觅食的核心,是一个所有能移动的生物体都要面临的永恒难题:是充分利用现有资源,还是去探索潜在的更优资源。这一问题极其根本,以至于大脑最初就是为解决这一问题而进化出来的,也从那时起,便不断围绕这个主题进行复杂化发展[4]。
在无模型探索(model-free exploration)中,动物通过随机搜索(对任何探索行为赋予相等的价值)和刺激-反应关联(即依赖习惯)来指导其行为,偶然发现有价值的信息[5]。
相比之下,基于模型的探索(model-based exploration)则依赖于动物对环境的认知模型及其目标导向性,在基于过去经验的环境心理模型中模拟各种行动,继而评估选项、预测结果。
由此,探索性觅食,从一个盲目的未知冒险,转变为一个不断优化的能量摄取的搜索过程。
04 内温性如何塑造神经认知?
鸟类和哺乳动物中观察到的神经认知适应现象支持了这样的假设:内温大脑在很大程度上是为了实现高效的基于模型的探索而进化的,以应对内温动物的能量需求。
这种进化的具体表现包括:鸟类和哺乳动物都独立演化出了更大的相对脑容量[6]和更高的神经元密度[7, 8],其信息处理能力和记忆容量也因此而显著提高。与体重相当的爬行动物相比,鸟类和哺乳动物的端脑(telencephalon)神经元数量分别多出平均17倍和9倍,小脑(cerebellum)神经元数量分别多出45倍和69倍[8]。
哺乳动物端脑的扩展与六层新皮层的出现密切相关,例如视觉信息通过拓扑和层级方式组织输入,组成视网膜定位编码(retinotopic coding);不同感官的感知信息在联合脑区整合,形成功能模块,例如边缘系统(limbic)、执行(executive)模块和运动前模块(premotor)。类似特征也在鸟类的端脑中有所体现,鸟类的超顶皮层(hyperpallium)也包含多个视觉场的拓扑映射[9]。这种结构可以实现快速详细的视觉分析,但需要比爬行动物视觉皮层中非视网膜定位编码(non-retinotopic coding)更多的神经元[10]。无论是鸟类或是哺乳动物,模块化脑区都变得越发复杂。
另外,鸟类和哺乳动物的端脑中还趋同地进化出了复杂行为协调区域——哺乳动物的前额叶皮层(Prefrontal Cortex, PFC)和鸟类的尾外侧顶皮层(Nidopallium Caudolaterale, NCL)[11]。这两个区域尽管解剖学上并非同源,但在功能上却高度相似。PFC和NCL均接收所有感官模态的输入信息、来自海马体的空间和记忆信息以及来自边缘系统的内部状态信息,将其输送到基底神经节和运动(前)结构,从而介导目标导向的行动[12],例如介导包括抑制和工作记忆等一系列执行功能。
除了皮层区域,鸟类和哺乳动物的海马体(hippocampus)的进化也更为趋同。虽然大多数脊椎动物的海马体都可以创建以外物环境为中心的空间地图(allocentric spatial map)[13]。然而,在鸟类和哺乳动物中,海马体的结构和连接性显然更复杂。它们扩大的海马体接收来自掌叶/皮层(pallium/cortex)的预处理信息,并拥有丰富的内在轴突侧支,使其无需大规模训练就能记住许多地点。鸟类和哺乳动物可能具有综合性的认知地图,包括与特定地点相关的“效价地图”(valence maps),以实现更快、更高效地编码记忆[14]。
除了在信息组织和编码上的相似性,鸟类和哺乳动物的大脑还独立演化出更复杂的基底神经节以实现更精细的行为模式控制[15]。鸟类单单基底神经节的神经元数量,就可达到甚至超过同等体型爬行动物整个端脑的神经元数量[7]。此外,为了更直接地控制运动,鸟类和哺乳动物都进化出了大脑中枢和下游兴奋性神经通路,这些通路对于灵巧性运动技能(如食物操作和提取性觅食)至关重要[16]。
小脑主要与运动控制和学习所需的预测性计算相关[17]。两栖类和有鳞目爬行动物的小脑较小、神经元数量较少,原鳄类(乌龟和主龙类)和合龙类动物的小脑的体积和神经元数量有所增加,鸟类和哺乳动物中最多[18]。此外,鸟类和哺乳动物进化出了端脑-小脑通路,其发育与端脑大小相关。虽然内温性动物的小脑的扩展,最初可能由复杂的陆地运动和直立姿势驱动的,以便在持续运动中实现高效呼吸。但越来越多的证据表明,小脑在各种认知功能中发挥着重要作用。
有效觅食的需求,推动了这些功能相似但独立进化的大脑结构进化。除了较为直接的感官运动控制外,鸟类和哺乳动物还发展出了更大规模更详细的认知地图。新进化的新皮层/小脑系统的一个关键特征是“离线”感官模拟,使动物能够在行动之前通过海马体认知地图评估不同选择。

▷图5.海马体认知地图功能相关细胞. 图源:Neuron
类似地,基底神经节无法区分感知模拟与真实,这可能促使动物基于能量效益更高的模拟场景进行行动决策。前额叶皮层(PFC)和尾外侧顶皮层(NCL)在权衡结果导向记忆与习惯性行为、决定是否抑制本能倾向或用新知识替代旧经验方面,起着至关重要的作用。这些自上而下的过程,有助于做出更明智的决策,并且被认为部分是为了增加热量摄入而进化得来的。
05 如何深入时间维度理解神经认知?
DeepTimeCognition团队将外温性与内温性动物的生存方式差异和神经解剖学证据相结合,发现了认知革命与内温性同时出现的进化现象;并通过认知神经科学研究揭示了大脑的演化过程和信息感知机制。关于外温性和内温性动物信息感知方式的深入研究,他们建议对位于关键系统发育节点的物种开展比较神经科学研究。

▷图6.深度时空解读神经认知的工作流程. 图源:Trends Cogn Sci
在羊膜动物(合弓纲)中,所有除内温性哺乳动物之外的物种都已灭绝。而爬行动物中,仍有众多存活物种,包括外温性动物(蜥蜴、蛇、坚蜥、龟和鳄鱼)和内温性动物(鸟类)。在爬行动物中,内温性大概出现在古爬形类中,该类群现存的代表物种包括鳄鱼(外温性)和鸟类(内温性)。鳄鱼大体保留了古爬形类的神经解剖结构,这些结构也是鳄鱼、非鸟类恐龙和鸟类的共同祖先特征。
古鸟类(Palaeognaths)是神经解剖学中发现的最原始的鸟类,与非鸟类伪鸟龙(Paraviandinosaurs)有许多共同点,它们也是鸟类中代谢率最低的,这可能反映了原始的能量调节方式。比较鳄鱼与古鸟类的认知能力,可以尽可能接近地还原古爬形类从外温性到早期完全高水平代谢外温性的演化过程。但迄今为止相研究很少,但现有结果认为古鸟类已经具有良好的社会认知能力。
研究除古爬形类以外的其他外温性爬行动物也很重要。在某些古爬形类中甚至更早期出现的神经认知适应性变化,或许促成了内温性的出现。例如,鳄鱼的如原始神经皮层前脑区(rudimentary NCL)和前脑-小脑投射等特殊结构,也在内温性动物中起着重要作用——这需要确认这些结构是否是古爬形类所特有的。
06 总结
伴随进化历程逐步提升的能量需求,体现在认知和神经解剖结构上。这种变化与从外温性向快速代谢的内温性转变趋势一致,并在鸟类和哺乳动物的进化路径上独立发生。认知能力支撑着生物体的生存,而代谢水平的提高促进了神经元数量的增加以及大脑新结构的出现,这些新结构反而又增强了认知效率,可以说——
动物如何“吃”和如何“想”,是一个紧密相连的过程。
DeepTimeCognition团队将外温性与内温性动物的生存方式差异和神经解剖学证据相结合,发现了认知革命与内温性同时出现的进化现象,并通过认知神经科学研究揭示了大脑的演化过程和信息感知机制。
以亿年为尺度回溯大脑的演化历程,不仅有助于我们探讨"智能"的本质,也能促使我们重新审视与自然界其他物种的关系。为了更好地理解这一认知转变,需要对处于关键系统发育节点的现存外温性动物和内温性动物进行全面的认知和神经解剖学比较研究。尽管当下研究任务艰巨,很多核心问题悬而未决,但开展此类研究的时机,或许已经成熟。
仍待追问:
现存的外温羊膜动物和内温羊膜动物,在认知方面的主要差异是什么?
外温性动物和内温性动物内部和之间,认知能力与大脑解剖结构有何关联?
能否在已灭绝物种中找出内温性动物特有的脑区相关性?
在内温性进化过程中,认知能力是如何逐步演化的?
参考文献:
[1] LYON P. Of what is “minimal cognition” the half-baked version? [J]. Adaptive Behavior, 2020, 28(6): 407-24. [2] NAGY K A. Food requirements of wild animals: predictive equations for free-living mammals, reptiles, and birds, F, 2001 [C]. [3] CAREAU V, KILLEN S S, METCALFE N B. Adding Fuel to the “Fire of Life”: Energy Budgets across Levels of Variation in Ectotherms and Endotherms [M]. Integrative Organismal Biology. 2014: 219-33. [4] CISEK P. Evolution of behavioural control from chordates to primates [J]. Philosophical Transactions of the Royal Society B: Biological Sciences, 2021, 377(1844): 20200522. [5] HILLS T T, TODD P M, LAZER D, et al. Exploration versus exploitation in space, mind, and society [J]. Trends in Cognitive Sciences, 2015, 19(1): 46-54. [6] SMAERS J B, ROTHMAN R S, HUDSON D R, et al. The evolution of mammalian brain size [J]. Sci Adv, 2021, 7(18). [7] OLKOWICZ S, KOCOUREK M, LUČAN R K, et al. Birds have primate-like numbers of neurons in the forebrain [J]. Proc Natl Acad Sci U S A, 2016, 113(26): 7255-60. [8] KVERKOVÁ K, MARHOUNOVÁ L, POLONYIOVÁ A, et al. The evolution of brain neuron numbers in amniotes [J]. Proc Natl Acad Sci U S A, 2022, 119(11): e2121624119. [9] BISCHOF H J, ECKMEIER D, KEARY N, et al. Multiple Visual Field Representations in the Visual Wulst of a Laterally Eyed Bird, the Zebra Finch (Taeniopygia guttata) [J]. PLoS One, 2016, 11(5): e0154927. [10] KNUDSEN E I. Evolution of neural processing for visual perception in vertebrates [J]. J Comp Neurol, 2020, 528(17): 2888-901. [11] MILLER E K, COHEN J D. An integrative theory of prefrontal cortex function [J]. Annu Rev Neurosci, 2001, 24: 167-202. [12] KERSTEN Y, MOLL F W, ERDLE S, et al. Input and Output Connections of the Crow Nidopallium Caudolaterale [J]. eneuro, 2024, 11(4): ENEURO.0098-24.2024. [13] MURRAY E A, WISE S P, GRAHAM K S. The evolution of memory systems: Ancestors, anatomy, and adaptations [M]. New York, NY, US: Oxford University Press, 2017. [14] MADISON F N, BINGMAN V P, SMULDERS T V, et al. A bird's eye view of the hippocampus beyond space: Behavioral, neuroanatomical, and neuroendocrine perspectives [J]. Horm Behav, 2024, 157: 105451. [15] FROST-NYLÉN J, THOMPSON W S, ROBERTSON B, et al. The Basal Ganglia Downstream Control of Action - An Evolutionarily Conserved Strategy [J]. Curr Neuropharmacol, 2024, 22(9): 1419-30. [16] TEN DonKELAAR H J. Evolution of Motor Systems: Corticospinal, Reticulospinal, Rubrospinal and Vestibulospinal Systems [M]//BINDER M D, HIROKAWA N, WINDHORST U. Encyclopedia of Neuroscience. Berlin, Heidelberg; Springer Berlin Heidelberg. 2009: 1248-54. [17] TANAKA H, ISHIKAWA T, LEE J, et al. The Cerebro-Cerebellum as a Locus of Forward Model: A Review [J]. Front Syst Neurosci, 2020, 14: 19. [18] HEUER K, TRAUT N, DE SOUSA A A, et al. Diversity and evolution of cerebellar folding in mammals [J]. Elife, 2023, 12.






 京公网安备 11011402013531号
京公网安备 11011402013531号